Phylum Protozoa
Protos — first, Zoan — animal, First animal phylum
Study of protozoans called Protozoology
Father of protozoology — Antony von Leeuwenhoek
Term Protozoa was assigned by Goldfuss
Protozoa belong to Kingdom Protista of Haeckel
Single-celled, solitary or colonial, eukaryotes
Important characters
Size ranges from 1µ to 5000µ
Aquatic — freshwater or marine forms, or endoparasites
Grade of organization- protoplasmic or subcellular
Locomotion by cilia, flagella or pseudopodia, or absent
Skeleton — may or may not be present; if present then either calcareous or siliceous. Foraminifers have external shell of CaCO3 ( ex. Polystomella), whereas Radiolarians have internal plates of silica
Osmoregulation by contractile vacuole in freshwater protozoans (isotonic to 0.8 % NaCl solution). C.V. absent in parasitic and most of the marine forms.
Symmetry — radial, bilateral or absent (asymmetrical)
Nutrition — Holozoic or holophytic.
Digestion intracellular within food vacuole. Reserve food material- Glycogen or Starchlike bodies
Respiration — exchange of gases occurs through plasma membrane
Excretion –excretory product NH3 (Ammonotelic). Excretion occurs through general body surface or plasmalemma.
Blood and nervous tissue absent. Sensory structure stigma (eyespot) present in Euglena
Reproduction — Asexual or Sexual. Asexual by budding, spore formation, binary fission or multiple fission. Sexual by Conjugation (Paramecium) or gamete formation (Plasmodium)
Euglena
Trypanosoma
Amoeba
Polystomella
Paramecium
Classification
Basis of classification — Locomotory structures
1. Sub Phylum Sarcomastigophora
(i) Super Class Mastigophora (Flagellata) —
Locomotory structures- flagella
Most primitive class
Binary fission –longitudinal
Two subdivisions- Phytomastigophora and Zoomastigophora
Phytomastigophora
Zoomastigophora
Green
Non-green
Chloroplast present
Chloroplast absent
Mostly free living
Mostly parasites
Reserve food material –starch (paramylon
bodies)
Reserve food material –Glycogen
Flagella- 1 to 2
Flagella 1 to many
ex. Euglena, Volvox, Noctiluca and Ceratium
Noctiluca –shows bioluminescence, called
Burning of sea
ex. Trypanosoma , Leishmania ,
Giardia, Trichomonas, Trichonympha,
Proterospongia, Mastigamoeba
T. brucei — digenetic parasite, primary host-cattle and secondary host — a fly; Disease-Nagana
T. cruzi — digenetic parasite, primary host- man and secondary host — dipteran fly; Disease- Chagas
T.evansi — digenetic parasite, primary host –horse and secondary host — Horsefly (Tabanus); Disease- Surra (skin disease)
Trichomonas — T. tenax aggrevates pyrrhoea, T. vaginalis causes leucorrhoea.
Leishmania donovani — a digenetic parasite, primary host- man and secondary host — Sandfly(Phlebotomus); Disease- Kala-azar(Dumdum fever)
L. tropica — causes Oriental sore or Skin Leishmaniasis
Giardia — a bilaterally symmetrical form, commonly called ‘Grand old man of intestine’; causes Diarrhoea
Trichonympha — It is a symbiotic protozoan in the gut of termites, for cellulose digestion.
(ii) Super Class Sarcodina (Rhizopoda)-
Locomotory structures-Pseudopodia ( false legs)
Free living or parasitic forms
Includes Heliozoans, foraminifers, radiolarians and amoebids
ex. Amoeba, Entamoeba, Polystomella, Arcella, Pelomyxa and Babesia (an RBC parasite, transmitted through tick and causes Texas fever in cattles)
Entamoeba histolytia — a monogenetic parasite in colon (large intestine) of man; causes Amoebiasis or Amoebic dysentry. Infective stage — Tetranucleated cyst. Mode of infection — contaminated food and water.
E. gingivalis (or buccalis) — a monogenetic parasite in the pockets, between teeth and gums; aggrevates pyrrhoea. Infective stage — Trophozoite. Mode of infection — Mouth to mouth kissing.
E. coli — a commensal of colon
2. Subphylum Ciliophora (Class Ciliata) –
The biggest and the most advanced class of protozoa
Binary fission — Transverse
Nuclei dimorphic, Micronucleus- reproductive and Macronucleus- vegetative in function.
ex. Paramecium, Vorticella and Rectal ciliates. Rectal ciliates –found in the rectum of frog and toad, include Balantidium, Nyctotherus and Opalina. Opalina has multinucleated condition. (micro- and macronuclei not differentiated).
Though the locomotory structures in Opalina are cilia, due to multinucleated condition, it is sometimes included in superclass Opalinata.
3. Subphylum Sporozoa -
Locomotory structures absent
Members most specialized for parasitic mode of life.
Exclusively endoparasites (contractile vacuole absent)
ex. Plasmodium, Monocystis and Eimeria
Monocystis- endoparasite in the seminal vesicle of earthworms, feeding on sperms
Eimeria- causes coccidiosis in poultry birds
4. Subphylum Cnidospora -
Spore formation throughout life
Spores contain polar capsule and coiled filament for attachment
Trophozoite — multinucleated
ex. Nosema, Myxidium (Nosema bombysis causes ‘Pebrine’ disease in silkworm, Bombyx mori).
The group Metazoa is divided into
Parazoa and Eumetazoa. Porifers
belong to parazoa as ectoderm in
these animals develops from vegetal
pole and the endoderm develops from
animal pole. In eumetazoans, it is just
reverse
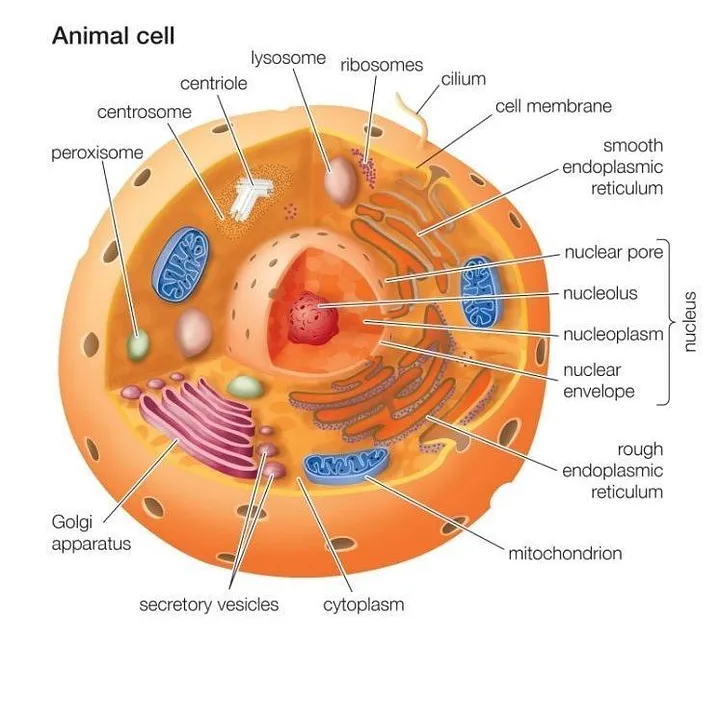
All animal cells are multicellular. They are eukaryotic cells. Animal cells are surrounded by a plasma membrane and it contains the nucleus and organelles that are membrane-bound. Unlike the eukaryotic cells of plants and fungi, animal cells do not have a cell wall. This feature was lost in the distant past by the single-celled organisms that gave rise to the kingdom Animalia.
Animal cells are of various sizes and have irregular shapes. Most of the cells size range between 1 and 100 micrometers and are visible only with the microscope. Trillions of cells are found in the human body. There are many different types of cells, approximately 210 distinct cell types in the adult human body.
The lack of a rigid cell wall allowed animals to develop a greater diversity of cell types, tissues, and organs. Specialized cells that formed nerves and muscles — tissues impossible for plants to evolve — gave these organisms mobility. The ability to move about by the use of specialized muscle tissues is a hallmark of the animal world, though a few animals, primarily sponges, do not possess differentiated tissues. Notably, protozoans locomote, but it is only via nonmuscular means, in effect, using cilia, flagella, and pseudopodia.
The animal kingdom is unique among eukaryotic organisms because most animal tissues are bound together in an extracellular matrix by a triple helix of protein known as collagen. Plant and fungal cells are bound together in tissues or aggregations by other molecules, such as pectin. The fact that no other organisms utilize collagen in this manner is one of the indications that all animals arose from a common unicellular ancestor. Bones, shells, spicules, and other hardened structures are formed when the collagen-containing extracellular matrix between animal cells becomes calcified.
Animals are a large and incredibly diverse group of organisms. Making up about three-quarters of the species on Earth, they run the gamut from corals and jellyfish to ants, whales, elephants, and, of course, humans. Being mobile has given animals, which are capable of sensing and responding to their environment, the flexibility to adopt many different modes of feeding, defense, and reproduction. Unlike plants, however, animals are unable to manufacture their own food, and therefore, are always directly or indirectly dependent on plant life.
Most animal cells are diploid, meaning that their chromosomes exist in homologous pairs. Different chromosomal ploidies are also, however, known to occasionally occur. The proliferation of animal cells occurs in a variety of ways. In instances of sexual reproduction, the cellular process of meiosis is first necessary so that haploid daughter cells, or gametes, can be produced. Two haploid cells then fuse to form a diploid zygote, which develops into a new organism as its cells divide and multiply.
The earliest fossil evidence of animals dates from the Vendian Period (650 to 544 million years ago), with coelenterate-type creatures that left traces of their soft bodies in shallow-water sediments. The first mass extinction ended that period, but during the Cambrian Period which followed, an explosion of new forms began the evolutionary radiation that produced most of the major groups, or phyla, known today. Vertebrates (animals with backbones) are not known to have occurred until the early Ordovician Period (505 to 438 million years ago).
Cells were discovered in 1665 by British scientist Robert Hooke who first observed them in his crude (by today’s standards) seventeenth century optical microscope. In fact, Hooke coined the term “cell”, in a biological context, when he described the microscopic structure of cork like a tiny, bare room or monk’s cell. Illustrated in Figure 2 is a pair of fibroblast deer skin cells that have been labeled with fluorescent probes and photographed in the microscope to reveal their internal structure. The nuclei are stained with a red probe, while the Golgi apparatus and microfilament actin network are stained green and blue, respectively. The microscope has been a fundamental tool in the field of cell biology and is often used to observe living cells in culture. Use the links below to obtain more detailed information about the various components that are found in animal cells.
Cell membrane
It is a semi-permeable barrier, allowing only a few molecules to move across it. Electron microscopic studies of cell membrane show the lipid bilayer model of the plasma membrane, it is also known as the fluid mosaic model. The cell membrane is made up of phospholipids which has polar(hydrophilic) heads and non-polar (hydrophobic) tails.
Cytoplasm
The fluid matrix that fills the cell is the cytoplasm. The cellular organelles are suspended in this matrix of the cytoplasm. This matrix maintains the pressure of the cell, ensures the cell doesn’t shrink or burst.
Nucleus
The nucleus is the house for most of the cells genetic material the DNA and RNA. The nucleus is surrounded by a porous membrane known as the nuclear membrane. The RNA moves in/out of the nucleus through these pores. Proteins needed by the nucleus enter through the nuclear pores. The RNA helps in protein synthesis through the transcription process. The nucleus controls the activity of the cell and is known as the control center. The nucleolus is the dark spot in the nucleus, and it is the location for ribosome formation.
Ribosomes
Ribosomes are the site for protein synthesis where the translation of the RNA takes place. As protein synthesis is very important to the cell, ribosomes are found in large number in all cells. Ribosomes are found freely suspended in the cytoplasm and also are attached to the endoplasmic reticulum.
Endoplasmic reticulum
ER is the transport system of the cell. It transports molecules that need certain changes and also molecules to their destination. ER is of two types, rough and smooth. ER bound to the ribosomes appear rough and is the rough endoplasmic reticulum; while the smooth ER does not have the ribosomes.
Lysosomes
It is the digestive system of the cell. They have digestive enzymes helps in breakdown the waste molecules and also help in detoxification of the cell. If the lysosomes were not membrane-bound the cell could not have used the destructive enzymes.
Centrosomes
It is located near the nucleus of the cell and is known as the ‘microtubule-organizing center’ of the cell. Microtubules are made in the centrosome. During mitosis the centrosome aids in dividing the cell and moving of the chromosome to the opposite sides of the cell.
Vacuoles
They are bound by a single membrane and small organelles. In many organisms, vacuoles are storage organelles. Vesicles are smaller vacuoles which function for transport in/out of the cell.
Golgi Bodies
Golgi bodies are the packaging center of the cell. The Golgi bodies modify the molecules from the rough ER by dividing them into smaller units with a membrane known as vesicles. They are flattened stacks of membrane-bound sacs.
Mitochondria
Mitochondria is the main energy source of the cell. They are called the powerhouse of the cell because energy(ATP) is created here. Mitochondria consists of the inner and outer membrane. It is spherical or rod-shaped organelle. It is an organelle which is independent as it has its own hereditary material.
Cilia and Flagella
Cilia and flagella are structurally identical structures. They are different based on the function they perform and their length. Cilia are short and are in large number per cell while flagella are longer and are fewer in number. They are organelles of movement. The flagellar motion is undulating and wave-like whereas the ciliary movement is power stroke and recovery stroke.
Endosomes and Endocytosis
Endosomes are membrane-bound vesicles, formed via a complex family of processes collectively known as endocytosis, and found in the cytoplasm of virtually every animal cell. The basic mechanism of endocytosis is the reverse of what occurs during exocytosis or cellular secretion. It involves the invagination (folding inward) of a cell’s plasma membrane to surround macromolecules or other matter diffusing through the extracellular fluid.
Intermediate Filaments
Intermediate filaments are a very broad class of fibrous proteins that play an important role as both structural and functional elements of the cytoskeleton. Ranging in size from 8 to 12 nanometers, intermediate filaments function as tension-bearing elements to help maintain cell shape and rigidity.
Microfilaments
Microfilaments are solid rods made of globular proteins called actin. These filaments are primarily structural in function and are an important component of the cytoskeleton.
Microtubules
These straight, hollow cylinders are found throughout the cytoplasm of all eukaryotic cells (prokaryotes don’t have them) and carry out a variety of functions, ranging from transport to structural support.
Peroxisomes
Microbodies are a diverse group of organelles that are found in the cytoplasm, roughly spherical and bound by a single membrane. There are several types of microbodies but peroxisomes are the most common.
In addition the optical and electron microscope, scientists are able to use a number of other techniques to probe the mysteries of the animal cell. Cells can be disassembled by chemical methods and their individual organelles and macromolecules isolated for study. The process of cell fractionation enables the scientist to prepare specific components, the mitochondria for example, in large quantities for investigations of their composition and functions. Using this approach, cell biologists have been able to assign various functions to specific locations within the cell. However, the era of fluorescent proteins has brought microscopy to the forefront of biology by enabling scientists to target living cells with highly localized probes for studies that don’t interfere with the delicate balance of life processes.
The main parts of a plant include:
Roots
Stem
Leaves
Flowers
Fruits
Roots
Roots are the important and underground part of a plant, which are collectively called the root system. They are the major part that anchors the plant firmly in the soil. They absorb water and minerals from the soil, synthesise plant growth regulators, and store reserve food material. The apical part of the root is covered by the root cap that protects the root apex.
The direct elongation of radicle leads to the formation of primary roots that grow inside the soil in dicots. It bears lateral roots that are known as secondary and tertiary roots.
In monocots, the primary root is replaced by a large number of roots because it is short-lived. In some plants such as Banyan tree, the roots arise from the parts of the plant and not from the radical. Such roots are known as adventitious roots.
Few plants that grow in swampy areas ave roots growing vertically upwards to get oxygen for respiration. Such roots are known as pneumatophores.
Stem
The stem is the part of the plant which is found above the ground. The bark of trees are brown in colour and younger stems are green in colour. It forms the basis of the shoot system and bears leaves, fruits and flowers. The region where the leaves arise is known as the node and the region between the nodes is known as the internode.
Stems arise from the plumule, vertically upwards to the ground. Initially, stems are usually weak and cannot stand straight. It eventually grows to become the toughest part of the plant called the trunk. The trunk is covered by a thick outer covering known as the bark. Overall stem provides a definite framework and structure to a plant, which later develops into a tree.
The stem provides support to the plant. They also protect the plant and help in vegetative propagation. A few underground stems such as potato and ginger are modified to store food.
The important functions of a stem include:
A stem carries out a number of functions essential for various processes such as photosynthesis.
Provides a definite framework and structure to a plant which later develops into a tree.
Support: Primary function of the stem is to hold up buds, flowers, leaves, and fruits to the plant. Along with the roots, a stem anchors the plants and helps them to stand upright and perpendicular to the ground.
Transportation: It is the part which transports water and minerals from the root and prepared food from leaves to other parts of the plant.
Storage: Stems are one of the storerooms of plants where the prepared food is stored in the form of starch. The stems of a few plants in the desert areas, such as Opuntia, get modified into thick, fleshy structures that store food and prevent excessive water loss due to transpiration.
Reproduction: Few stems help in reproduction through vegetative propagation and also help to bear flowers and to produce fruits.
Guards: Protects Xylem and phloem allowing them to perform their functions. The stem tendrils are spirally coiled and help the plant to climb support. The axillary buds also get modified into thorns that protect the plant from grazing animals.
The stems of a few plants in the desert areas, such as Opuntia, get modified into thick, fleshy structures that store food and prevent excessive water loss due to transpiration.
Leaves
Leaves are the most important part of a plant. They contain chlorophyll that helps the plants to prepare their food using sunlight, carbon dioxide and water. A leaf consists of three main parts- petiole, leaf base and lamina.
The petiole keeps the leaf blade exposed to wind and cools the leaf.
The leaf base is a protruding part of a leaf.
The lamina of the leaf contains veins and veinlets that provide rigidity to the leaf blade and help in the transport of mineral nutrients.
Primarily, leaves have three main functions:
Photosynthesis: Green leaves prepare food for plants by using water and carbon dioxide in the presence of sunlight. This process is called photosynthesis.
Transpiration: Other than photosynthesis, leaves play a crucial role in the removal of excess of water from plants through tiny pores called stomata. This is the process of transpiration.
Reproduction: Leaves of some plants helps in reproduction also. For e.g. leaves of Bryophyllum give rise to a new Bryophyllum plant.
Other Parts of Plants
The other parts of a plant include flowers and fruits.
Flowers
Flowers are the most beautiful and colourful part of a plant. They are the reproductive part of a plant. A flower has four major parts, namely,
Petals: It is the colourful part of a flower which attracts insects and birds.
Sepals: Sepals are green leafy parts present under petals and protect the flower buds from damage.
Stamens: This is the male part of the flower consisting of anther and filament.
Pistil: This is the female part of the flower consisting of stigma, style, and ovary.
Fruits
Fruits are the main features of a flowering plant. It is a matured ovary that develops after fertilisation. Some fruits are developed without fertilization and are known as Parthenocarpy or parthenocarpic fruits.
Thus, we see how different parts of a plant help in the growth and development of a plant. All the plant parts are beneficial and work in coordination with each other.
Excretion is the process whereby an organism eliminates metabolic wastes and unwanted chemicals from its system. Metabolism is the sum total of all the chemical reactions occurring in the cells and body. Some products of these metabolic reactions are toxic and so must be processed or eliminated from the body. Others are simply materials that are present in excess and so must be eliminated as waste. The process of excretion is quite different to defecation, which is the removal of undigested food wastes from the gut. However, the gut of many animals also has a role in excretion as some materials may be excreted into the gut and eliminated with the faeces. In insects most excretory products are excreted into the gut lumen and eliminated along with faecal matter. Excretion is also important in eliminating excess water and other unwanted chemicals that may be ingested and enter the body fluids, such as plant poisons and excess salts.One of the main functions of excretion is to remove excess nitrogen. Nitrogen enters the diet in the form of amino acids, nucleic acids and certain salts. One of the main products of excretion in aquatic organisms is ammonia. Ammonia contains nitrogen and is a small molecule which dissolves readily in water. This allows it to be easily excreted into the surrounding water. However, this becomes a problem for terrestrial organisms. Ammonia is toxic to cells and so must be quickly ejected from the body, however, being water-soluble it is typically ejected in solution, which requires water. The mammalian solution is to convert the ammonia into a less toxic substance called urea. This conversion takes place in the liver: the ammonia produced by cells enters the bloodstream where the liver removes it, converts it into urea which again enters the bloodstream to be excreted by the kidneys. Being less toxic, the urea can be temporarily stored and excreted in a concentrated solution, requiring less water.Birds and reptiles have a better water-conserving system; they excrete uric acid (or urate salts). Uric acid is not readily soluble in water and is of low toxicity and so can be excreted with very little water. The dry excreta of birds is a mixture of faecal matter and uric acid crystals and when water is scarce birds can produce very dry excreta.Arthropods, including insects, have adopted similar solutions. Woodlice, which are not insects but crustaceans, are only partially adapted to terrestrial conditions, preferring moist habitats, but they do excrete ammonia. Interestingly they can vent off ammonia gas, rather than relying on the wastage of water to remove the ammonia in solution. Insects are better adapted to dry conditions, although aquatic insects and some insect larvae excrete ammonia, most terrestrial forms excrete uric acid (or salts of uric acid called urates, such as ammonium urate).If one considers how small an insect is and how rapidly a small drop of water may evaporate, then one realises that insects have outstanding water-conserving systems. Bedbugs (Rhodnius) can survive for weeks without ingesting any water! Some insects can tolerate extremely dry conditions and may excrete uric acid as a dry crystalline powder, along with bone-dry faeces! Insects generally produce only trace amounts of urea. Malpighian tubules The main excretory organ of the insect is the Malpighian tubule. Insects contain anything from 2 to 150 or more Malpighian tubules depending on the genus. Malpighian tubules are tubular outgrowths of the gut. They typically develop as pouches emerging from the junction between the midgut and the hindgut, though there actual final position varies — they may be attached to the midgut, hindgut or the midgut-hindgut junction as is the case with our ant above.Each Malpighian tubule is a blind-ending tube whose lumen is continuous with the lumen of the gut. Each consists of a single layer of epithelial cells, forming the tubule wall, enclosed by an elastic membrane (basement membrane — a fibrous and porous protein mesh). In most insects there is a thin layer of striated muscle around this membrane. Typically muscle cells spiral around the distal end (the end furthest from the gut) of the tubule, causing it to twist and turn in gentle writhing movements as the muscles contract. The proximal end (near the gut) may be coated in circular and longitudinal muscle fibres, giving rise to peristalsis or squeezing movements which empty the contents of the tubule into the gut. In some cases, such as in caterpillars, the Malpighian tubules on each side (3 on each side in this case) empty into a small bladder, which then empties into the gut. In this case only the bladder may be muscular and its lumen is lined by cuticle (suggesting that the bladder is an extension of the hindgut).The tubules do not just hang around in the air! The body cavity of the insect is filled with a fluid, usually colourless, called haemolymph. This fluid bathes the organs and tissues and is circulated around the insect body. The tubules are also typically loosely or firmly anchored in place by the tracheae which attach to them.The twisting and turning of the Malpighain tubules presumably keeps them in contact with fresh haemolymph (perhaps by circulating the heamolymph around the tubule). Metabolic wastes and other unwanted chemicals that entered the insect system pass into the haemolymph, or are excreted into the haemolymph by the cells. These include nitrogenous waste and plant toxins such as alkaloids. It is the job of the Malpighian tubules to keep the haemolymph cleansed of these wastes — they remove wastes from the haemolymph and then excrete them into the gut lumen.Outside the muscle layer is a ‘peritoneal covering’ of cells with embedded tracheoles, which carry oxygen to the Malpighian tubules which their mitochondria use to generate the needed ATP by aerobic respiration. How do Malpighain tubules work? Waste materials and excess water pass from the haemolymph into the Malpighain tubules, by crossing the epithelial wall of these blind-ended tubes. Recent evidence shows that these cells contain pumps, proteins called proton-secreting V-ATPase. These proteins use energy in the form of ATP (see respiration) to pump protons into the lumen of the Malpighian tubule. Protons are positively charged and to maintain charge balance the removal of protons from the epithelial cells, into the tubule lumen, is balanced by the inward movement of potassium ions, which move from the haemolymph, into the epithelial cells and then out into the tubule lumen also. The diagram below shows a section through a segment of a Malpighian tubule. The epithelial cells have microvilli (fingerlike projections) projecting into the tubule lumen and are rich in mitochondria (green stripy rods) which produce the ATP required by the pumps. A model of how ion transport across the epithelium is thought to take place is illustrated.
The detailed structure of the cell at top right has been simplified to illustrate some of the transport mechanisms. The V-ATPase is shown as the orange circle pumping protons (H+) into the tubule lumen. Removal of the protons from the epithelial cell makes the cytoplasm more negatively charged and also sets up a concentration gradient (that is an electrochemical gradient is established) and this attracts positive ions, such as sodium (Na+) and potassium (K+) into the cell from the haemolymph. The influx of these positive ions drags in negative chloride ions to balance the charge. These ions move across the cytoplasm of the cell, the so-called transcellular pathway. Note the potassium-chloride and sodium-chloride symporters, the proton-potassium and proton-sodium antiporters and the ion channels.The flux of ions across the epithelial cell also draws across water, by osmosis. This probably takes place largely by the paracellular pathway, that is between the epithelial cells. Sugars and amino acids are swept along by the water into the tubule lumen. Since these materials are useful they will be reabsorbed later downstream.Other small molecules (small enough to cross the basement membrane) will also move into the tubule through this pathway. The transport of a substance which depends directly on ATP, such as the pumping of the protons in the Malpighian tubule, is called active transport. The transport of the other ions and water is passive (by facilitated diffusion) in of itself, but is dependent on proton transport and so indirectly dependent on ATP. This mode of transport is called secondary active transport, e.g. the transport of potassium.In dry conditions many insects can produce a very concentrated urine, indeed one that is ‘bone-dry’. However, many insects ingest large quantities of water when feeding, such as blood sucking insects, and in this instance the rate of fluid-flow through the Malpighian tubules increases a thousandfold or more. Indeed, the rate of fluid transport in these tubules is said to be higher, gram for gram, than any other tissue. Two hormones, released into the haemolymph, can stimulate Malpighian tubules to rapidly increase their rate of fluid transport: 5HT (5-hydroxytryptamine) and a peptide hormone. Increased excretion is triggered by an increase in uric acid following a meal, which presumably triggers the release of the diuretic (urine-producing) hormones.Of course, not all the fluid transported through the tubules is excreted. The proximal (basal or lower or downstream) sections of the tubules, along with the hindgut (especially the rectum) reabsorb some of the water, depending on need, and other useful substances, such as certain ions, sugars and amino acids, so as to produce a final urine of the ‘desired’ concentration. It is in this proximal or lower part of the tubule that uric acid is transported into the tubule, against a concentration gradient, and precipitates as crystals, e.g. of insoluble potassium urate as the urate combines with the high potassium content of the tubule lumen. In some insects these crystals can be seen filling the lumens of the proximal ends of the tubules. Presumably, peristalsis then moves these crystals along into the gut. Potassium and some of the chloride are recovered in this way, producing a urine high in sodium.
Some small organic molecules are also actively transported into the tubule lumen by the transcellular pathway, including alkaloids (plant compounds which may be toxic to the insect).Uric acid, mostly in the form of negatively charged urate ions, is also actively transported by the transcellular pathway, though the exact mechanism is not well understood. This urate transport occurs in the proximal tubule and the urate combines with the potassium transported into the tubule to form insoluble potassium urate crystals. These crystals form roughly spherical concretions in the tubule lumen. The microvilli in the proximal tubule seem to undergo a cycle of elongation, as the urate concretions form, and retraction as the lumen fills up with urate waiting to be transported into the gut.Once in the gut, remaining water may be reabsorbed as needed and the remaining urate excreted with the faeces, or separately. The midgut is divided from the hindgut by the pyloric sphincter and when this sphincter is closed the hindgut receives only the contents of the Malpighian tubules.The mechanism of excretion demonstrated by the Malpighian tubule is one largely dependant on ‘secretion’ of unwanted materials, such as urate and excess sodium. This contrasts with the mammalian kidney which relies on ultrafiltration (filtration through microscopic pores), which removes most materials from the blood except large proteins and cells, followed by reabsorption of what the body needs to keep, such as sugars and amino acids. However, there is some filtration in the Malpighian tubule, namely the influx of materials through the paracellular pathway, having filtered across the basement membrane. Sugars and amino acids filtered in this way are then reabsorbed, as in the mammalian case. Similarly, there is some secretion in the mammalian kidney, for example the secretion of protons and ammonium in acid-base balance and the secretion of some drugs such as penicillin. However, the emphasis is different with the Malpighian tubule relying more on secretion, the mammalian kidney on filtration. Other mechanisms of excretion Some insects, such as the silverfish, springtails and aphids have no Malpighian tubules. Stick insects may have three types of Malpighian tubules. Clearly much remains to be learnt about excretion in insects. In addition to excretion by Malpighian tubules, insects often exhibit storage excretion in which waste materials are sequestered safely and kept inside special storage cells. For example, the fat body may contain urate cells which accumulate urate crystals throughout the life of the insect. pH regulation and other functions of Malpighian tubules The main function of Malpighian tubules may be the elimination of nitrogenous waste, but hand in hand with this comes the task of water conservation (eliminating waste whilst conserving water when necessary) or osmoregulation — regulating water content of the insect body and also regulation of ion balance. Considering their involvement in cleansing body fluids of unwanted materials it is not surprising that excretory organs typically have major roles also in regulating acid-base balance. Enzymes only work within a narrow range of acidity or pH and so an organism has to excrete excess acid or excess base to maintain the correct pH of its body fluids. Malpighian tubules also have a role in acid-base balance. The V-ATPase actively excretes protons and hence excess acid (an acid is a chemical which generates protons in solution as the protons are the true source of acidity).Calcium is also excreted in large quantities by the Malpighian tubules of some insects. Generally, some of the Malpighian tubules, or one specific segment of the tubules, takes on this function. These tubules often become distended as they fill with calcium salt crystals. Some insects make use of this calcium in the construction of their burrows or larval cases, such as the helical calcium carbonate shells of some spittlebug (Ptyelus) larvae.Finally, the Malpighian tubules of some insects may assume a glandular function in the secretion of silk.